暑假開始,終於有更多時間翻譯喜歡的文獻了~~~
第三彈終於翻譯完成,希望有進步
原文在此:(太長了所以分段,整體即爲網址)
http://www.sciencemag.org/content/32...1339.abstract?
maxtoshow=&HITS=10&hits=10&RESULTFORMAT=&andorexacttitle=and&andorexacttitleabs=and&fulltext=
Molecular+and+Evolutionary+History+of+Melanism+in+North+American+Gray+Wolves
&andorexactfulltext=and&searchid=1&FIRSTINDEX=0&resourcetype=HWCIT
===============================================
Molecular and Evolutionary History of Melanism in North American Gray Wolves
北美洲灰狼黑變病的分子和進化曆史
作者群:Tovi M. Anderson, Bridgett M. vonHoldt, Sophie l. Candille, Marco Musiani, Claudia Greco, Daniel R. Stahler, Douglas W. Smith, Badri Padhukasahasram, Ettore Randi, Jennifer A. Leonard, Carlos D. Bustamante, Elaine A. Ostrander, Hua Tang, Robert K. Wayne, Gregory S. Barsh
摘要:親緣關係緊密的物種的形態學多樣性是進化和適應的一個基本形式。黑色素皮質激素受體1(Melanocortin 1 receptor, Mc1r)基因的突變有助於魚、鳥和許多哺乳類野生種群獲得多樣的毛色。然而,灰狼(Canis lupus)的黑變病是由一種與衆不同的黑素腎上腺皮質激素(melanocortin)途徑形成,即K位點,其編碼作爲Mc1r供選擇配合基的β-防禦素蛋白(beta-defensin protein)。我們指出北美狼的黑化K位點突變來源於曾經和家犬的雜交,這個突變的頻率在森林生境很高,同時我們提出了這個分子標簽(molecular signature)存在積極選擇。相同的突變也在郊狼(Canis latrans)和意大利狼中造成黑變病,因此我們的結論證明了家養物種的特征選擇(traits selected)如何影響他們野生近親的形態學多樣性。
點擊以顯示隱藏內容
毛色和生境的相似性(correspondence)常常歸因於自然選擇(natural selection),但很少有分子水平(molecular level)的證據。北美灰狼的種群中,北美西部森林和開放生境(open habitats)毛色頻數分析不同,這些生境包括德奈利國家公園(Denali National Park)、阿拉斯加洲的基奈半島(the Kenai Peninsula)和許多加拿大北極地區。這些差異,尤其是高緯度跟隨北美凍原馴鹿(barren-ground caribou)遷徙到其繁殖地的苔原狼,和終年居住在鄰近北方森林並以不遷徙的獵物爲食的狼之間的,最爲引人注目。苔原生境中,深色狼非常稀少,而在西南方漸變群到森林地區,頻率卻逐漸增高(圖1A)。深毛色對淺毛色可能的選擇包括捕獵時的隱蔽和/或基因多效性(pleiotropy)的間接效應,但仍有疑團未解開,因爲還有未被識別的潛在基因存在。
在許多脊椎動物中,自然的顔色變異受Agouti-黑色素皮質激素受體1(Mc1r)途徑控制,雙配體受體(ligand receptor pair)調節位於皮膚、毛發或羽毛的黑素細胞(melanocyte)産生色素——紅/黃色素或棕/黑色素——的數量和類型。功能獲得性Mc1r突變會導致一些家養和實驗動物以及一些鳥類、齧齒類和犬科野生種群的黑變病,這是被廣泛認可的。最近,我們發現家犬中的色素類型開關包含了一個黑素腎上腺皮質激素途徑的額外組分,即K位點編碼的β-防禦素蛋白CBD103。
加拿大狼群的毛色基因是複雜的,表現型從白到灰到黑,同時這些表型還被因年齡增大而灰化的獨立事件所混淆(圖1B)。然而,在小群建群者是被從加拿大引入的黃石國家公園裏,灰和黑色毛皮呈孟德爾形質(Mendelian trait)分離。我們調查Agouti基因、Mc1r和CBD103的分子變異,並在北美狼中識別了一些Mc1r和Agouti基因多態性。然而,這些基因功能影響都無法預測,而且也與黑色毛皮沒有聯係。與此相反,在一個來自黃石、有14個成員、共三代的群體中,我們觀察到黑毛和K位點標記【連鎖LOD分數的讓步比對數(LOD)爲4.21,估計最高的可能重組分數(θ)爲0(圖1C)】之間完全的共分離(co-segregation),這是未知的並且依賴帶有Agouti和Mc1r基因的染色體。
在狗中,祖先傳下的CBD103等位基因(Ky)令Agouti和Mc1r基因正常作用,而其中3個堿基對(base pair, bp)缺失突變(CBD103ΔG23或KB)抑制Agouti基因作用,導致表現型爲黑毛的顯性遺傳。我們在來自黃石的104只黑狼中觀察到有102只擁有同樣的3個堿基對缺失突變,而此突變在來自加拿大極地的9只黑狼中均存在。相反地,在來自黃石的120只灰狼中均沒有CBD103ΔG23,來自加拿大極地的22只白狼也均無(表1)。我們同樣在10只來自加拿大極地的灰狼中發現有6只存在CBD103ΔG23,表明灰色毛皮可由CBD103ΔG23改良agouti表現型導致(毛發既含有淡黃色色素,也含有真黑素),也可由次要因素如年長會淡化毛發中的色素沈積(pigmentation)以致只剩下真黑色素導致。【附加黃石狼種群係譜研究和其中少量的Mc1r變異,表明基因型爲ky/ky的黑色黃石狼可能因在取樣時表現型不明確而誤分類】 CBD103ΔG23等位基因頻率在苔原和森林的狼總體估計分別爲0.02和0.19,相應的黑毛表現型頻率爲33%和64%(圖1A)。
爲調查黑化K等位基因的進化曆史,我們依序排列分散在CBD103基因大約150kb的範圍內的8個單拷貝非編碼DNA片段,這些片段來自32只極地狼和15只沒有親緣關係的黃石狼,以及12只家犬:6只ky/ky(秋田犬akita、巴辛吉犬basenji、拳師犬boxer、鬥牛犬bulldog、杜賓犬Doberman pinscher和大丹犬great dane)、6只KB/KB(卷毛尋回犬curly-coated retriever、大麥町犬Dalmatian、大丹犬、拉布拉多尋回犬Labrador retriever、貴賓犬poodle和葡萄牙水獵犬Portuguese water dog)。我們在所有犬科中識別出52種等位基因多態性(狼有36種),並估計每個單體型結構(haplotype structure)(圖2B)。所有狼的擴增子(amplicon)中,多態性比率是每510 bp一個單核苷酸多態性(single-nucleotide polymorphism, SNP)(Watterson估值,qW = 1.96 × 10−3),這相似於拳師犬和灰狼(580 bp之1)或和郊狼(420 bp之1)的全基因多態性組測量值。然而,根據我們關於K位點基因型的數據進行分類,以及含KB基因的染色體和CBD103距離很近,表現型與CBD103相似或相同,上升達到或超過那些觀察到的含ky染色體位於橫跨位點任一邊的75kb(圖2A)。這種模式,以及核苷酸差異的同功性(π,圖S1),同樣反應在單體型多樣性的顯著差異,KB(22之8)和ky(72之59)染色體(c2 = 14.2, P < 0.001)。再加上毛色和生境的相關性,低多樣性和高頻率相結合,表明KB基因在北美的森林狼中被正選擇。
總的說來,K位點周圍150 kb的基因的連鎖不平衡(linkage disequilibrium, LD)模式相似於不同品種家犬之間的對比,有相對地小型單倍域(haplotype blocks),這其中包括曆史上沒有重組迹象的大約4kb的CBD103核心序列(圖2C)。然而,北極狼KB和ky等位基因的不同進化曆史很明顯,而SNP(圖2B)被描繪成單體型分叉圖(圖2D),這突出顯示了狼KB單體型中心區大約60kb的多態性缺乏。這個特征以及KB和ky染色體相應的差異,顯示出擴展單體型純合度(extended haplotype homozygosity, EHH)統計數值,這兩種隨機選擇的染色體保留了從CBD103逐漸增加距離統一性。正如圖2、E和F所示,相比狼的ky染色體,狼的KB的EHH的分布相當廣,然而在狗之中這兩種染色體的EHH分布幾乎相同。再加上額外的基因組SNP數據分析,這些選擇性掃描(selective sweep)觀察表明KB已升至高頻率。
正如同黑狗和黑化的狼一樣,CBD103ΔG23也在67只郊狼(6黑61灰,表1)中被證明和毛色相關。這些發現顯示出三種可能的進化曆史。第一,這種3bp的缺失突變可能相當古老,古老到它是在一百萬年前的犬科祖先中發生的,那時郊狼和狼還沒有分開。第二,這種3bp的缺失突變可能發生得更近,在其中一個物種中,然後經由趨中雜交傳播至其他物種。第三,這種3bp的缺失突變可能是一個突變熱點,分別發生在郊狼、狼和狗中。爲區分這些可能性,我們查明並將郊狼的單體型(6個KB和18個ky)和北美狼、狗相比較。
這三種犬科動物的單體型多樣性和在狼中觀察到的相似,而且KB有明顯的較低多樣性(40之中有15是獨特的),相比ky(102之中有66是獨特的)染色體(c2 = 9.7, P = 0.003)。15種獨特的KB單體型中,一個在3只郊狼和6只狗中被觀察到,一個在2只郊狼和12只狼中被觀察到(圖3A)。然而,66種獨特的ky單體型沒有一種同時在兩種或以上不同物種中被觀察到。重建全部150kb區域的係統發生網絡和現存KB與ky染色體間的曆史重組(圖2B中的箭頭),但缺少適當的方式用於推斷在重組情況下的精確基因係譜。然而,在CBD103的4kb核心區域(圖2C),根據18個核心區單體型建立的簡單鄰位相連進化樹(neighbor-joining tree)標示出142個(94狼,24狗和24郊狼)染色體(圖3B)。在這個進化樹上,所有的KB染色體形成2-單體型簇,反之剩下的16種單體型(這些代表所有的ky染色體)更加分散。此外,許多ky染色體按物種形成一簇(狗的12分之9和狼的72分之44),和KB染色體不同。這具有鮮明對比的係統發生圖表明KB突變是發生在一個物種裏,接著再通過種間雜交(interspecific hybridization)分散到狗、狼和郊狼之中。【來自郊狼的24個ky單體型不像來自狼或狗的那樣彼此相似(圖3B),這與他們和其他犬科動物雜交的曆史相吻合。】
要深入了解在狗和狼中産生的K位點變異,我們根據來自狼、狗和兩者集合的ky和KB染色體的CBD103累積距離,估計最近共同祖先(the most recent common ancestor, TMRCA)的合並時間。我們使用了分子鍾(molecular clock)的方法,排列個別全150kb區域擴增子的數據(圖2),假設突變發生在狼和狗的所有位點有一相同固定比率,混合重組和統計的影響。CBD103附近,TMRCA估計所有KB子集(圖3C)接近0,因爲在這個範圍內多態性很小或沒有(圖3A)。然而,無論基因型如何,在離CBD103更遠的位點(10到50kb),狗染色體與狗和狼染色體合集相似。這說明狗中的KB足夠古老到和ky染色體進行廣泛的重組,這個重組曆史還包括狗和狼的雜交。但是,在相同的10到50kb的範圍內,TMRCA估計狼的KB染色體子集不超過狗的KB染色體子集(或狗和狼KB染色體合集),這說明KB是由狗向北美狼引入的,或者反之亦然。
KB從狗至北美狼引入的論點同樣被地理和生態因素支持。KB廣泛分布在家犬中,包括起源在亞洲和非洲的古老品種。然而在狼中,北美之外只有意大利有報道過黑變病,在那裏近期存在狼和自由放養的狗之間的雜交,且有分子和/或形態學的證據。實際上,我們同樣分析了來自意大利亞平甯山脈的22個樣本,在7只黑狼中發現6只有KB(包括一只狼狗混血),但在15只灰狼中沒有一只存在KB。相比之下,10個KB/ky和10個ky/ky基因型的北美狼的全基因組SNP分析顯示,沒有證據表明狗和狼之間近期存在雜交。
狗是在15,000至40,000年前從東亞的灰狼馴化而來,我們估計KB突變至少已有46,886年的曆史(95%置信限度:12,779到121,182年);因此,我們無法區分KB到底是在馴化之前還是之後出現的。然而,如果KB出現在舊世界未馴化的狼中,我們的數據表明它一定曾在基因庫(gene pool)中遺失,之後又在北美狼中重新獲得,可能來自12,000至14,000年前跟隨人類度過白令海峽(Bering Strait)的北美原住狗(圖3D)。
美國的狼面臨嚴峻的威脅,有時是捕殺引起,有時來自混種雜交,例如在北美五大湖的情況。然而,北美灰狼中明顯的KB位點選擇表明基因多樣性——被人類保存在家犬中的——可能在野生狼種群中興盛。由於可用的苔原生境因開發和/或全球變暖而減少,KB突變的頻率可能在發展到更北的地方。因此,原本在家養動物中發生並選擇的突變引入野生種群,這種基因多樣性的引入可能、具有諷刺性地提供了一種令野生種群更加適應不斷變化的環境的機制。種間雜交也在其他家養動物和植物中被發現。我們的研究結果意味著,家養生物中的突變型可能在野外環境中存活並豐富野生種群的基因遺産。
表一:CBD103在狼和郊狼中的分布
點擊以顯示隱藏內容
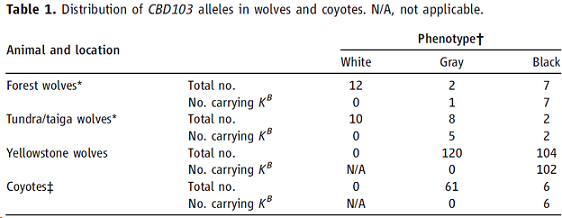
*森林和苔原/針葉林的狼來自加拿大極地地區(圖1A)。在森林和苔原/針葉林中全部深色(灰或黑)狼的頻率分別爲62%和7%,這種基因型分配不代表種群頻率。所有森林和苔原/針葉林的狼中,帶有KB的基因型爲KB/ky;在黃石國家公園的種群中,10只是KB/KB、92只是KB/ky。
†表現型的絕對的指定,由采集的樣品定義,沒有完全獲得正常毛色的變化範圍,如圖1B中的文字描述。
‡灰色郊狼來自內布拉斯加州(Nebraska)或西弗吉尼亞州(West Virginia);黑色郊狼來自明尼蘇達州(Minnesota)或西弗吉尼亞州。
圖1:黑變病和K位點基因型在北美洲灰狼中的分布。
點擊以顯示隱藏內容
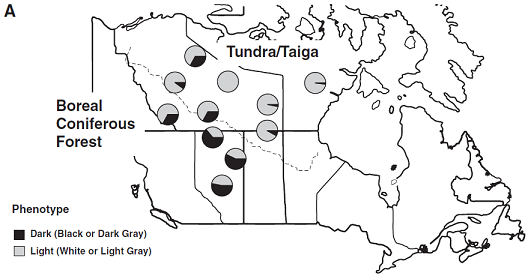
(A) 這裏所使用的加拿大樣本的位置和毛色表現型。

(B) 年齡相關的灰化,以及由表現型猜想得出的基因型。每一組照片顯示同一個體在不同年齡(10個月和10歲),並在10歲時發現漸增的灰色毛皮,表明在KB/ky個體中真黑素被稀釋(左邊一組圖),在ky/ky個體中真黑素和色素都被稀釋(右邊一組圖)。(照片由印第安納州Battle Ground, Wolf Park的Monty Sloan提供。)
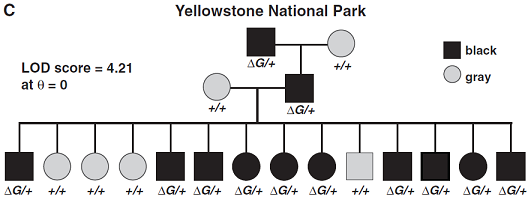
(C) 來自黃石公園的Leopold pack的三代純種狼的KB和黑毛的共分離。ΔG表示顯性KB等位基因,+表示野生型等位基因ky。
圖2:K位點的多態性和單體型結構,北美灰狼【(A)到(E),1KB/KB,20KB/ky和26ky/ky】和家犬【(F),6KB/KB和6ky/ky】。
點擊以顯示隱藏內容
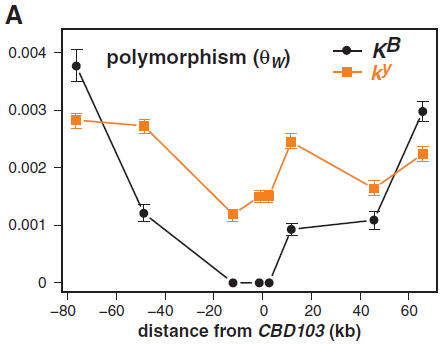
(A) 根據到CBD103的距離計算的多態性(ΘW,±SD)。
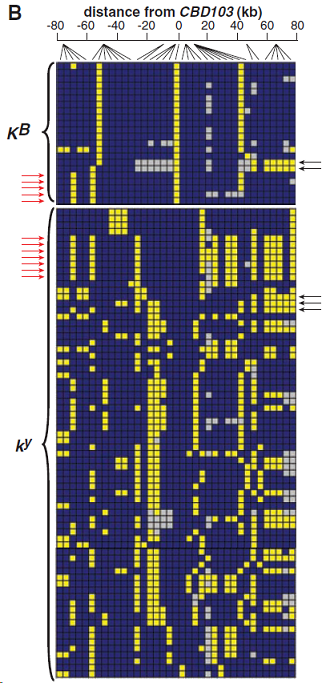
(B) 在36個SNP上推斷的狼的單體型結構;每一行代表一條KB或ky染色體;藍色和黃色方塊分別代表主要和次要等位基因;灰色方塊代表缺失數據。紅色和黑色箭頭分別在5’和3’末端指出代表KB和ky染色體存在重組曆史的例子。
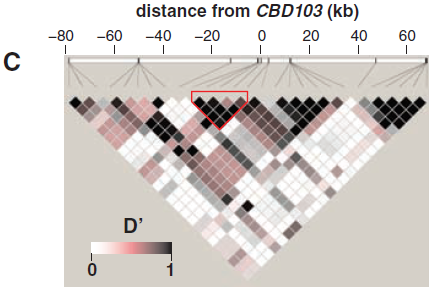
(C) 所有狼染色體的成對的LD值(以D'表示);紅色輪廓表示一個沒有重組曆史的核心區域(同圖3)。
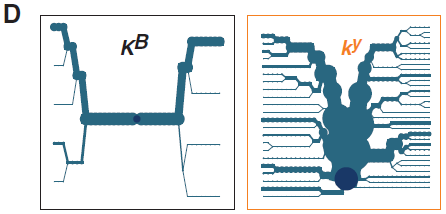
(D) KB或ky染色體的單體型分歧圖,深藍色圓點代表CBD103,分支代表單體型分歧,分支的線條粗細和染色體數目成比例。
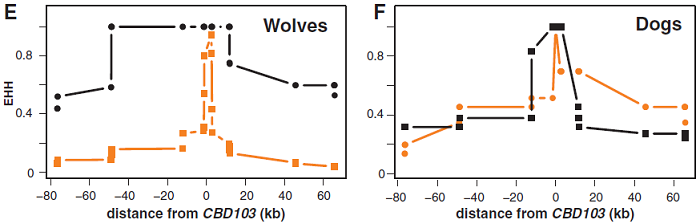
(E和F) 根據到CBD103ΔG23距離計算狼(E)和狗(F)的KB或ky染色體的EHH值。
圖3:犬科動物中K位點的進化關係和曆史。
點擊以顯示隱藏內容
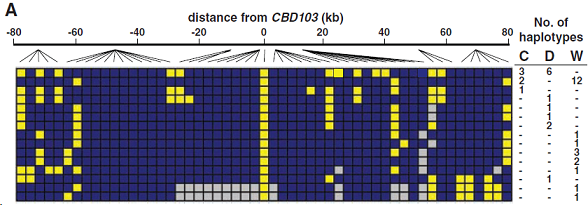
(A) 似狼犬科動物(wolflike canid)的KB單體型結構,基於52個SNP。每一行代表在郊狼(C),狗(D)和狼(W)中發現的KB單體型,在右側列出他們各自的頻率,顔色和圖2B中的意義相同。
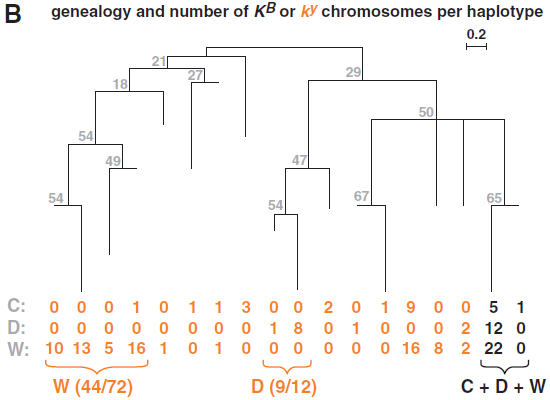
(B) 推斷出核心區域(圖2C)單體型的係譜關係【來自500拷貝的自展值(bootstrap value)標示在分支旁】。每條分支代表18種不同單體型之一,每種單體型的染色體數目下標明物種。
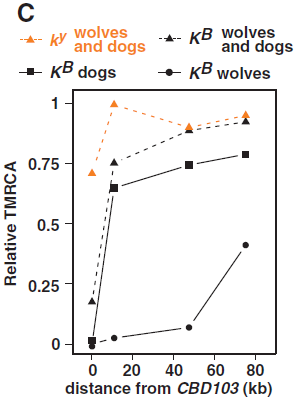
(C) TMRCA估計表明依據分子鍾計算的染色體子集,表示所有似狼犬科動物的部分分歧時間。函數代表有親緣關係的TMRCA隨與CBD103距離增大而增加,可能由於祖先的雜交和重組。
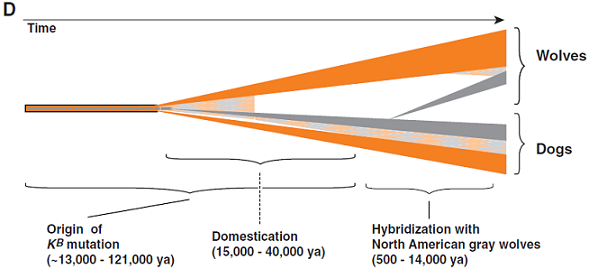
(D) K位點在狗和狼中演化的時間軸,橙色部分表示祖先的ky染色體,灰色部分表示衍生的KB染色體,橙灰相間的圖案表示重組染色體。ky至KB的突變和馴化時期重疊或在那之前,但令KB進入北美灰狼基因庫的趨中雜交發生在更近的時候。
===============================================
文中KB、ky和CBD103ΔG23的B、y和ΔG23均爲上標
文和圖都較多,而且文還沒有分副標題,一直在想要不要圖文鑲嵌式(?)
結果還是分開來排版比較方便……
依然是相當分子水平的文獻,細節相當複雜~~
這個研究曾被新聞報道過: 先有狼 還是 先有狗呢!?
可是人家研究主要結論是家養動物的基因突變如何影響了野生種群的遺傳多樣性
而且還一直在強調狗確實是由東亞狼演化而來
跟狼和狗出現的先後到底有啥關係?
明明新聞內文問題不大,這題目到底取些什麽亂七八糟的東西?





 回覆時引用此篇文章
回覆時引用此篇文章